Groei-gekoppelde enzym engineering door manipulatie van redox cofactor regeneratie
Groei-gekoppelde enzym engineering door manipulatie van redox cofactor regeneratie

Een steeds breder scala aan chemicaliën wordt nu geproduceerd uit hernieuwbare grondstoffen via biotechnologische middelen. Bioprocessen zijn sterk afhankelijk van enzymatische katalyse voor de efficiënte productie van deze verbindingen. Ervoor zorgen dat enzymen optimaal presteren in hun vereiste omgeving is daarom van groot belang voor duurzame productie.
Eigenschappen zoals substraatspecificiteit, katalytische snelheid en (thermo)stabiliteit behoren tot de verschillende kritische factoren die moeten worden geoptimaliseerd voor efficiënte enzymgestuurde bioprocessen. Deze optimalisatie kan tijdrovend, kostbaar en uitdagend zijn, en daarom zijn effectieve en kostenefficiënte manieren waarschijnlijk voor groei-gekoppelde selectie, technische enzymen met wenselijke kenmerken zeer gewild.
De voordelen van het gebruik van groeigekoppelde selectie als een vorm van enzymselectie, waarbij de activiteit van een enzym wordt gekoppeld aan de groei van een cel. Deze methode kan worden gebruikt als een selectiestrategie met hoge doorvoer en kan worden bereikt door ervoor te zorgen dat de groei afhankelijk is van productsynthese door het doelenzym of door de activiteit van het enzym te koppelen aan de globale energietoestand van de cel. Synthetische biologie kan worden gebruikt om stammen te ontwikkelen die geschikt zijn voor groeigekoppelde selectie, en recente ontwikkelingen hebben zich gericht op technische stammen met een tekort aan de geoxideerde of verminderde toestanden van redoxcofactorparen, die kunnen dienen als enzymtechnologieplatforms. Het gebruik van deze platforms kan de ontwikkeling van verbeterde biokatalysatoren en bioprocessen versnellen.
Voordelen van groeikoppeling via cofactor auxotrofie
De voordelen van het gebruik van cofactorauxotrofie als selectiemethode voor het engineeren van enzymen die betrokken zijn bij de biosynthese van bepaalde chemicaliën, zoals lipiden, biobrandstoffen, gassen, organische oplosmiddelen of polymere verbindingen. Deze aanpak biedt verschillende unieke voordelen, waaronder de mogelijkheid om onafhankelijk van het substraat of product van belang te selecteren voor het gewenste product, waardoor het gemakkelijker wordt om verbeterde enzymatische activiteit te detecteren en een uitlezing te bieden voor het detecteren van verbeterde enzymatische activiteit. Bovendien is het gebruik van cofactor-auxotrofen als groeikoppelingsplatforms gunstig omdat de alomtegenwoordige aard van redoxcofactoren in het microbiële metabolisme betekent dat engineeringstrategieën lateraal overdraagbaar kunnen zijn naar andere microben van belang, en enzymen kunnen direct worden gemanipuleerd in de omgeving van de gewenste microbiële gastheer. (weergegeven als figuur 1)

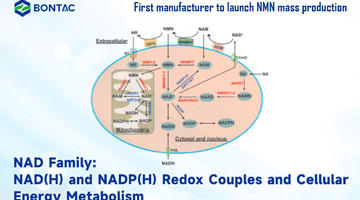
Figuur 1: Vergelijking van verschillende veelgebruikte screening/selectietechnieken voor enzymtechnologie
Mutanten met een tekort aan NADH-oxidatie
De oxidatie van NADH in E. coli kan via twee routes plaatsvinden, afhankelijk van de beschikbaarheid van zuurstof. Onder aerobe omstandigheden wordt NADH voornamelijk geoxideerd door ademhaling om ATP te genereren, terwijl het onder anaërobe omstandigheden kan worden geoxideerd via fermentatieroutes om lactaat en ethanol te produceren. Gemuteerde stammen van E. coli, niet in staat om gemengde fermentatieroutes te gebruiken voor NADH-oxidatie tijdens anaerobe groei, zijn gebruikt om NADH-oxidatieroutes aan te sturen voor de anaërobe synthese van verschillende chemicaliën, zoals 2-methylpropan-1-ol, 2,3-butaandiol, 1-butanol en L-alanine. Deze gemuteerde stammen zijn ook gebruikt om enzymen te engineeren door gebruik te maken van vergelijkbare redoxprincipes, wat resulteert in verbeterde varianten. De resulterende stammen kunnen worden gebruikt om andere NAD(P)H-afhankelijke enzymen en routes te engineeren.
Mutanten met een tekort aan NAD+ reductie
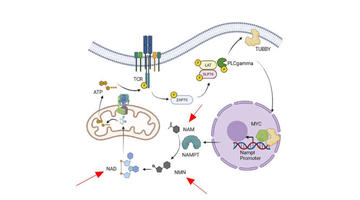
Wenk et al. creëerden een E. coli-stam door het dihydrolipoyldehydrogenase (lpd) -gen te verwijderen, wat ertoe leidde dat de stam niet in staat was om reducerend vermogen (NADH en NADPH) uit het acetaatmetabolisme te genereren vanwege de afwezigheid van pyruvaatdehydrogenase-activiteit. Dit zorgde ervoor dat de stam auxotrofie vertoonde voor het verminderen van het vermogen wanneer het aeroob werd gekweekt op acetaat als enige koolstofbron. (weergegeven als figuur 2) De stam was in staat om op acetaat te groeien wanneer het werd aangevuld met bovenste glycolytische substraten of bij het tot expressie brengen van NAD + -afhankelijke formate-, ethanol- of methanoldehydrogenasen met hun respectieve substraten. De stam werd niet gebruikt voor enzymtechnologie en werd alleen beoordeeld op redoxcofactor auxotrofie.

Figuur 2: Centraal metabolisme
Mutanten met een tekort aan NADPH-oxidatie
Er zijn twee verschillende strategieën voor het induceren van NADP + auxotrofie in E. coli, afhankelijk van het engineeren van de glycolytische route om NADPH te overproduceren. De eerste strategie omvat het verwijderen van het inheemse gapA-gen en het tot expressie brengen van een heterologe NADP + -afhankelijk GAPDH-enzym, terwijl de tweede strategie bestaat uit het omleiden van de koolstofflux via de pentosefosfaatroute. De resulterende stammen zijn niet in staat om op glucose te groeien, maar vertonen groei in verschillende omstandigheden, waarbij de eerste stam onder anaërobe omstandigheden wordt gekweekt en de tweede onder aerobe omstandigheden met glycerol als substraat. Deze stammen worden gebruikt voor het engineeren van enzymen met verbeterde eigenschappen, waaronder substraatspecificiteit, katalytische activiteit en thermostabiliteit.
Mutanten met een tekort aan NADP+ reductie
Drie verschillende bacteriën, E. coli, P. putida en C. glutamicum, zijn ontworpen om NADPH-auxotroop te zijn, wat betekent dat ze exogene NADPH nodig hebben voor groei. In het geval van E. coli en C. glutamicum werden centrale metabole enzymen uitgeschakeld om NADP + -reductie te voorkomen wanneer glucose als koolstofbron werd verstrekt, terwijl in P. putida CRISPR / nCas9-geassisteerde engineering werd gebruikt om sets doelgenen sequentieel te verstoren om hun betrokkenheid bij het redoxmetabolisme te begrijpen. De NADPH-auxotrofe stammen werden vervolgens gebruikt voor groei-gekoppelde enzym engineering op basis van cofactor specificiteit. Een enkele ronde mutagenese met behulp van E. coli leverde de meest efficiënte en specifieke NADP+-afhankelijke formatedehydrogenase tot nu toe op, terwijl P. putida en C. glutamicum de eerste stammen van hun soort vertegenwoordigen die voor dit type engineering kunnen worden gebruikt.
Mutanten met een tekort aan NMN+ reductie
Er werd een groeigekoppeld selectiesysteem ontwikkeld om cofactorcycli en groei te koppelen op basis van een tekort aan NMN+-reductie. De E. coli SHuffle-stam werd gebruikt, die deleties draagt in twee genen die betrokken zijn bij de productie van gereduceerd glutathion. Een NMNH-afhankelijk glutathionreductase werd ontwikkeld door middel van rationele mutagenese om het proces te koppelen aan NMN + / NMNH-cycli, en een NMN + -afhankelijke glucosedehydrogenase werd gebruikt om NMN + / NMNH-cofactorcycli en dus groei te ondersteunen. De studie paste ook voor het eerst niet-canonieke redoxcofactorauxotrofie toe voor groeigekoppelde enzymtechnologie, wat resulteerde in een thermostabiele fosfietdehydrogenasevariant met verbeterde katalytische efficiëntie en temporele stabiliteit in vitro. Het werk biedt een nuttige stam voor groei-gekoppelde enzym engineering, afhankelijk van NMN + / NMNH-cycli en auxotroof voor de verminderde toestand van een niet-canonieke redoxcofactor.
Het potentieel van groeikoppeling via redoxcofactoren werkte als een krachtig hulpmiddel om biokatalysatoren te engineeren, met name in de context van duurzame bioproductie. Hoewel er aanzienlijke vooruitgang is geboekt bij het genereren van redoxcofactor auxotrofe stammen, suggereert de verklaring dat er nog steeds verschillende onontgonnen wegen voor onderzoek zijn. Een daarvan betreft het engineeren van cofactor auxotrofe stammen van andere organismen dan E. coli, wat nieuwe mogelijkheden zou kunnen openen voor groei-gekoppelde enzymtechnologie.
Over het algemeen suggereert het dat het gebruik van op redoxcofactoren gebaseerde groeikoppeling een belangrijke kans biedt voor technische biokatalyse, met name voor de productie van producten die niet kunnen voldoen aan de vereisten van andere high-throughput enzymtechnologiebenaderingen. Het benadrukt de noodzaak om meer veelzijdige en efficiënte biokatalysatoren te ontwikkelen voor duurzame bioproductie en onderstreept het belang van voortgezet onderzoek op dit gebied.
Referentie: Jochem R. Nielsen a, Ruud A. Weusthuis b, Wei E. Huang a, Growth-coupled enzyme engineering through manipulation of redox cofactor regeneration, Biotechnology Advances, 2023.